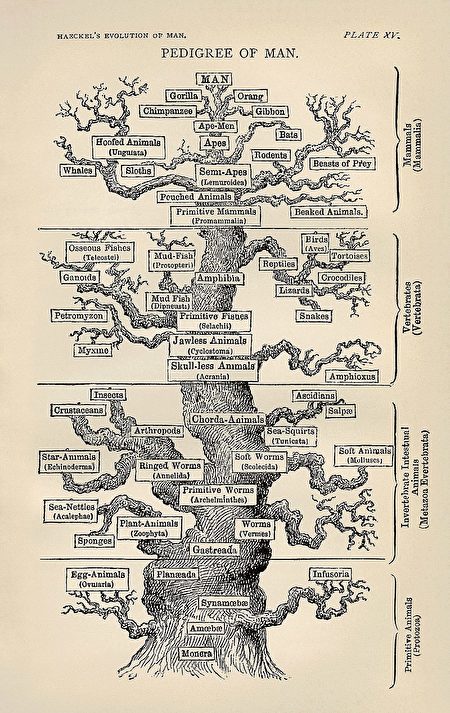
海克尔1866年绘制的错误的人类系统进化树(The tree of life as seen by Haeckel in The Evolution of Man. By Ernst Haeckel – First version from en.wikipedia; description page was here. Later versions derived from this scan, from the American Philosophical Society Museum. Public Domain)
1966年德国生物学家威利‧亨尼格(Willi Hennig, 1913—1976)出版的《系统发育系统学》(Phylogenetic Systematics),成为系统生物学的基础。在这本书中,亨尼格坦率承认特别重要的“排序”的任务是如何艰难和复杂。²³²
当人们对物种的排序一筹莫展的时候,恰逢现代分子生物学的发展给人们带来了大量新的基因数据。刚开始看到这些数据的人,就像进入一个琳琅满目的糖果商店,感到十分兴奋。兴奋之余,人们就开始通过采用各种各样的基因数据和方法,组建各种各样的系统发生树。
人们通常会根据不同物种基因的差异程度构建进化树。进化树的每个节点(Node)代表共同祖先或导致物种分化的事件。树枝代表进化的分支,也称为演化支(clade),而树叶通常表示一个具体的物种。建构进化树时,可以使用有根法(rooted)来找到假定的共同祖先,或使用无根法(unrooted)无需假定共同祖先。
目前存在多种建构进化树的方法,包括邻接法(Neighbor joining, NJ)、似然法(Likelihood)、简约法(Parsimony)和贝叶斯法(Bayesian)。由于数据或方法的限制,有时候可能出现一个节点分出多个演化支的情况。
需要注意的是,这只是对建构进化树过程的简要描述,具体的方法和应用可能更加复杂且多样。
1990年,美国微生物学家卡尔‧沃斯(Carl Woese, 1928—2012)、德国植物学家和微生物学家奥托‧坎德勒(Otto Kandler, 1920—2017)与美国微生物学家马克‧L‧惠利斯(Mark L. Wheelis)提出了一个由古细菌、细菌和真核生物三个系统组成的“生命之树”。²³³
后来,人们根据生物体中广泛分布的核糖体RNA(rRNA)的小亚基16S(18S) rRNA的基因重建假设的生命进化树。²³⁴
由生物学家和数据库开发人员等合作开发的、提供所假定的物种演化的在线资源TimeTree,初版于2007年发布。
2015年TimeTree包含2,274篇发表的文章涉及的50,632个物种,以螺旋的形式呈现²³⁵。同年9月,发布了包含更多新物种数据的开放生命之树²³⁶。2016年发布的一种新版本,纳入了更多的基因数据²³⁷。2022年,第五版发布,包含4,075篇已发表的文章涉及到的137,306个物种,是迄今为止规模最大的版本。²³⁸
尽管“进化树”在整个发展过程中耗费了众多科学工作者的时间和精力,貌似越来越繁荣壮大,但其实它根本上是错误的,存在着与生俱来的理论缺陷。
4.2 假想的“节点”
构建“系统发生树”的一个大前提是,假定一个物种和另一个物种之间存在“共同祖先”。它是以现有的基因学证据或形态学证据推论过去发生的亲缘关系事件。
“(做)系统发育分析最基本的假设是,一棵树上的所有基因序列都是同源的,即来自一个共同的祖先。无论是否同源,都会用这个假设来比对基因序列。所有系统树都会根据该对齐方式构建。然而,如果这些序列实际上不是来自共同祖先,那么这棵树将毫无意义,而且很可能会产生误导。”²³⁹(The most basic assumption of phylogenetic analysis is that all the sequences on a tree are homologous, that is, descended from a common ancestor. Alignment programs will align sequences, homologous or not. All tree-building programs will make a tree from that alignment. However, if the sequences are not actually descended from a common ancestor, the tree will be meaningless and may quite well be misleading. )
也就是说,“系统进化树”上不同物种之间的亲缘关系都是假想出来的,也就是说,树的“节点”是假想的、虚构的、根本就不存在的,那些看似枝繁叶茂的参天大树整个都是虚构出来的,根本就无法站立。
而且,目前未发现可以用来印证不同物种之间存在“共同祖先”的证据。考古学、地质学、分子生物学的大量证据表明,地球上的生命起源不是像达尔文所描述的那种单一、线性、漫长演化的模式,而且也没有先后顺序;相反,地球上生命具有多次爆发→灭绝的重复循环的特点。
很多科学研究都不支持物种演化假说,例如寒武纪突然集中产生的20至35个不同门类生物的化石²⁴⁰,《人类进化》的DNA条形码快照研究发现地球上十分之九的物种都是在同一个时期(10万到20万年前)出现的²⁴¹,犹他州的峡谷国家公园出土的3亿年前脊椎骨骼遗骸的化石²⁴²,还有许许多多早在进化假说所假定的人类出现之前的、更久远年代就已经存在的人类的脚印²⁴³,都构成了显明的反对达尔文所臆想的“物种进化”的铁证。
所以,地球上的不同物种的产生,没有时间上的先后关系,而是几乎同时、平行产生的,根本就没有共同祖先,这些节点也根本不存在。所谓的参天大树是不存在的,实际上可能只是一个个矮小的“灌木”丛。
不同物种基因之间的相关性以及同一物种内的变异度可能是存在的,但这些都不能证明不同物种之间的“宏进化”,即不同节点之间的物种的跳跃。
4.3 虚构的“目录”
由于系统进化树一定要把地球上林林总总的生物按照不同类别、假定的物种产生时间做一个排序,就牵扯到分类的问题。
在传统分类当中,各个类群都有着比较清晰的鉴定特征。例如哺乳动物的特征包括胎生哺乳、有毛发、恒温等等,鸟类的特征包括有羽毛、具有喙、卵生等等。
但是,分类方法是一种人为的产物,在等级划分和命名上往往有非常强的主观性。
打个比方,我们用电脑管理文件的时候,一个个的文件是客观个体,但是目录结构是人为划分的,每个目录里面摆放什么文件也是人定的。一个文件,既可以放到目录A中,也可以放到目录B中,也就是文件与文件之间的位置关系是人定的。这类似于一个个物种是存在的,但是物种之间的亲缘关系是人为假想的。
一些生物的奇特的结构和功能也带来许多不解之谜,比如章鱼。
章鱼具有非常奇特的身体构造,它有3颗心脏、8个腕足和极其复杂的大脑及神经系统²⁴⁴。章鱼有5亿个神经元,除了主要的中央大脑,章鱼的每条腕足都有一小群控制运动的神经细胞,相当于还有8个独立的迷你大脑²⁴⁵。
章鱼具有一些超凡脱俗的能力:可以解决谜题、改变自己的形状和颜色,甚至编辑自己的基因。与人类的基因组相比,章鱼的蛋白质编码基因多出了33,000个,而且几乎每个蛋白质编码基因中,都存在广泛的保守的腺苷到肌苷(A-to-I)mRNA编辑位点。与之对比,人类只有1%~3%的蛋白质编码mRNA具有类似位点²⁴⁶。
更令人意外的是,被归类到软体动物中的头足类的章鱼,在5亿年前就出现在地球上了。它的这种复杂而又独特的结构和功能让科学家十分困惑:在系统进化树的哪里去摆放它的位置呢?这真是一个巨大的难题。
正当科学家倍感困惑的时候,2018年,英国白金汉大学天体生物学家、美国密歇根州生物物理中心的一批学者与来自澳洲的学者大胆地在《生物物理学和分子生物学进展》(Progress in Biophysics and Molecular Biology)上发表论文提出,章鱼可能是寒武纪大爆发时,附着在彗星上降落在地球的生物。²⁴⁷
让科学家倍感困惑的奇特的章鱼。(amretsunique/Shutterstock)
关于“章鱼到底是怎么来的”这个问题的答案,我们在后面的章节会继续探讨,但是,科学家这个大胆的猜想,其实反映了人们对地球上生物无比复杂、实在无法理解的一种大胆思索。至少这种思维打破了达尔文只把地球当作一个封闭的体系来看待的那种狭隘、片面地解释生命起源的模式。
其实每个物种是独立存在的,物种之间被强加的进化关系,也就是被“系统进化树”虚构出来的“目录”路径,根本就不存在。
4.4 拓扑学错误
系统发育分析是一项复杂繁琐的任务,需要大量的数据处理²⁴⁸。例如,比较不同位点的测序分类群、处理缺失数据以及对所有位点的分析进行复制和比较。在系统进化树的分析过程中会出现不可避免的拓扑学错误。
拓扑学是一门研究空间形状和连接性质的数学学科,就像孩子们玩的“橡皮泥”,俗称“橡皮泥几何”。它主要关注的是物体的变形、拉伸和弯曲。可以把它想像成探索物体之间如何连接在一起的方式。
拓扑学的理论在进化树中得到应用。系统进化树的拓扑学错误可以用一个比喻来解释,想像你要绘制一个家族树,表示家族成员之间的亲属关系。如果你在绘制家族树时,错误地将某些人归类在不正确的分支上,或者遗漏了某些人的亲属关系,那么这就是家族树的拓扑错误。这个错误可能会导致家族树不能准确地反映真实的亲属关系。
同样的,系统进化树的拓扑学错误指的是在建构生物进化关系的树状图时,出现了类似的错误,使得物种之间的关系图不准确,将不同物种之间错误连接或者错误地将某些物种归类在同一分支上。
早在1989年,享誉世界的美国国家科学院院士、美国遗传学会主席迈克尔‧林奇(Michael Lynch, 1951—)在《进化》(Evolution)期刊上发表文章指出:“进行亲缘树分析时可能出现很多问题。如果物种之间的分歧只是由于随机漂移和突变引起,六个或更多物种之间距离的系统发育树几乎总是包含拓扑错误。……进一步考虑到物种之间不断的选择差异,将导致完全消除准确建树的可能性,当然也包括系统发育树的分支点。”(Even with enormous amounts of data, phylogenetic trees fitted to the distances among six or more species will almost always contain topological errors if the mode of divergence has been random drift and mutation. ……Moreover, on closer consideration, it appears that the realization of the assumption of constant selection differentials between species would eliminate completely the possibility of identifying the branch points in a phylogeny.)²⁴⁹
简单总结,即使在最简单的中性突变的模型下,当通过输入种种数据来重建系统发生(生物进化)的关系时,当数据超过六个物种的时候,就已经无法重建物种之间的系统发生(生物进化)关系了。
也就是说,简单的树只要稍微大一点,六个或更多物种之间无法保证不出错,更不用说复杂的树的模型了。因为现有进化论模型无法应对生物复杂性,好比物理学里的多体问题或量子力学里的测不准原理。上面的分析其实是推导出了分子生物学里的测不准原理。
可是为什么这么多的大数据依然无法建立起一棵稳固的系统进化树呢?
究其根本原因,是因为达尔文“物种进化”是错误的假说。用达尔文的错误的“物种进化”思想试图去理解生命的起源、解释生命基因的大数据,就像盲人摸象,永远也无法获知大象的全貌;又像海中捞月,永远只能是幻想。
所以人们在开始“建树”时往往热情高涨,可是实际应用起来,往往是困难重重。比如越是行业内、领域内的精英人士,越是心知肚明一件事──“系统进化树”有这样一个与生俱来的“拓扑”错误,直接否定了这棵树的可行性。很多的建树的文章,往往只是为了获取学位所走的过场,而并非为了获取什么真正的知识。这个缺陷打中的是“进化树”的死穴,它是一棵根本不可能立起来的树,因为它必倒无疑。
五、自我矛盾的“系统进化树”
当人们刚刚获得大量基因组数据的时候,如获至宝,以为找到了一个很好的用来描绘“生命之树”的工具。可是经过一段时间的应用之后,人们越来越发现,基因数据出现的自相矛盾的情况越来越多。
正如1998年美国《科学》期刊的科普作家伊丽莎白‧彭尼西(Elizabeth Pennisi)在《科学》期刊上发表的标题为“基因组数据撼动生命之树”的文章指出的那样:“这些基因数据给人们带来的混乱远远多于启发。”²⁵⁰
5.1 不同基因,不同结果
首先,进化论支持者通常假定所有生物都起源于一个祖先,试图寻找在生物基因里面“进化上保守”(也就是通常稳定不变)的基因序列。而到底选择哪个基因来构建“进化树”,可谓是一个“见仁见智”的问题。
美国分子遗传学家费尔德曼(Rorbert Feldman)接受彭尼西的采访时表示:“用什么样的基因,就会得到什么样的进化位置。”²⁵¹
举个例子,细菌研究领域里,美国微生物学家和生物物理学家卡尔‧理查德‧沃斯(Carl Richard Woese, 1928—2012)提出来,根据细菌核糖体(Ribosome)的小亚基的保守成分16S rRNA基因序列来确定细菌的相似度,以此来确定不同细菌的亲缘关系。²⁵²
可是人们后来发现,不同物种之间不光是垂直的传宗接代的关系,不同物种之间还存在不可思议的基因水平转移(horizontal gene transfer),指生物将遗传物质传递给其它细胞而非其子代的过程的现象,导致进化树的结构更加混乱。所以彭尼西在《科学》文章中还指出:“但新测序的微生物基因组以及与酵母等真核生物基因组的比较,使这一清晰的图景变得混乱,引发了对所有生命分类的怀疑。”²⁵³
5.2 混乱的“家谱”辈分
另一个大问题是,同样的数据,采用不同的算法,会得到不同的系统发生树的结果。最后经常是“公说公有理、婆说婆有理”,造成长期争论。
大家都熟悉族谱,在族谱里,同一世代的人被放在同一水平位置上,兄弟姊妹是一代,父母是一代,祖父母是一代。
在系统进化树中,如果用不同的基因或不同的分类方法,可能出现很不一样的辈分关系。
一个典型的例子是学术界对裸子植物亲缘关系的争论,纷扰近30年却仍未有定论。裸子植物可分为苏铁、银杏、柏类、松类和买麻藤类,与被子植物一起组成种子植物。过去的观点认为买麻藤与被子植物亲缘较近,但基于DNA研究的演化生物学已推翻了这一观点。现在有多个猜想,其中一派认为买麻藤与松类最相近,另一派认为买麻藤与柏类最相近,还有一派认为买麻藤是其它裸子植物的姊妹群,还有另一派认为买麻藤是其它种子植物的姊妹群。²⁵⁴
结果出现了不同观点的人各执一词、谁也说服不了谁的局面,五种不同的“进化树”同时共存。每一种进化树中,不同种的植物所处的辈分,一会儿是孙子、一会儿是爷爷,就好比同一个人在一个家族里面,用一种方法分析是爷爷辈的,用另一种方法分析又被鉴定为孙子辈,真是让人啼笑皆非。
与之类似,山雀的谱系也是五花八门。虽然将多个基因视为单个进化单位是一个很诱人的假设,但许多问题可能导致出现完全不同的系统进化树。²⁵⁵
5.3 “以不变应万变”的生存伎俩
“进化论”支持者在构建“进化树”的过程中,为了自圆其说而添补漏洞,去解释达尔文提出的原版、线性、单一的“进化论”所无法解释的现象,有目的地引入了平行进化(parallel evolution)²⁵⁶、发散进化(divergent evolution)²⁵⁷、趋同进化(convergent evolution)²⁵⁸等等新概念,不断扩展原版进化论所涵盖不了的“进化”模式。
比如,为了解释为什么章鱼的眼睛具有与人的眼睛相同的复杂结构,就提出了所谓“平行进化”;再比如为了解释为什么狗和狼的共同祖先能分别进化出狗和狼这么不同的物种,就提出了所谓“发散进化”。
可是,殊不知,这些被设计出来的进化新模式,早已经打破了原版达尔文的框架,原版的“进化假说”已经不知不觉嬗变为一个奇奇怪怪的理论体系。而进化论支持者不管生物如何千变万化,有一个中心思想是永恒不变的,那就是永远都要在“进化”的框子里面演变,永远都将其归入“进化树”的形形色色的变化模式之中。
这就是“系统进化树”为了应对各种难题和困扰所采取的“以不变应万变”的生存伎俩。
六、自相矛盾的“分子钟”
在建树的过程中,人们通常会用到分子钟(Molecular clock)技术,分子钟也叫基因钟、演化钟,也就是根据某些基因的突变率推断两个或多个生物在演化历史上分离的时间,因此,分子钟已成为系统进化分析的重要组成部分和重要工具。²⁵⁹
经常被用来判定生物亲缘关系远近的蛋白质的基因有:细胞色素C、组蛋白、血红蛋白和纤维蛋白肽等。根据不同的蛋白质来计算,会有一个不同的分子时钟。
例如,细胞色素C(Cytochrome C)位于细胞内的线粒体之中,是生物氧化过程中的电子传递体。其作用原理为在酶存在的情况下,对组织的氧化、还原有迅速的酶促作用。进化论学者通常认为细胞色素C是一种进化上保守的分子,所以它经常被用来分析生物之间的亲缘关系。
微生物学家迈克尔‧道顿(Michael Denton)博士在《进化论:陷入危机的理论》(Evolution: A Theory in Crisis)中指出²⁶⁰:
人和鲤鱼的血红蛋白序列相差50%,细胞色素C仅相差13%。如果进化假说是正确的,血红蛋白分子钟在以比细胞色素C更快的速度“进化”。也就是说,有两种以不同速率滴答作响的分子钟。
不仅如此,由于有数百个不同的蛋白质家族,每个家族都表现出其独特的种间变异程度,有些大于血红蛋白,有些远小于细胞色素,因此不仅仅是两个时钟,而是数百个蛋白质家族,每个家族都被视为以其独特且高度特异性的速率运行。
毫无疑问,不同蛋白质的不同“进化速率”已经成为人们描绘“系统进化树”一个不可逾越的分子壁垒。因此,分子钟自首次使用以来的几十年间,从最早的、简单的同速模型,已经变得越来越复杂,建立模型和方法也越来越复杂。选择什么样的分子钟模型来建树,绝不是一项简单的任务。²⁶¹
问题的复杂性不仅仅如此,使用任何一种所谓的“进化上保守”的蛋白质分子来构建“系统进化树”,都会出现与之前按照“进化论”所假设的前提相矛盾的结果。
道顿还指出:“尽管细胞色素C序列在不同的陆生脊椎动物中有所不同,但它们与鱼类的序列是等距的。在分子水平上,没有鱼类→两栖动物→爬行动物→哺乳动物的进化轨迹。因此,传统上一直被认为介于鱼类和其它陆生脊椎动物之间的两栖动物,在分子方面与鱼类的距离与任何爬行动物或哺乳动物的距离一样远!对于那些熟悉脊椎动物进化的人来说,这个结果确实令人吃惊。”²⁶²
没有迹象表明,被进化论认为的“高等”生物比“低等”生物“进化”得更多。
原核生物细菌的细胞色素C的氨基酸序列,与酵母、小麦、蚕蛾、鸽子和马等真核生物,都具有接近的相似度(64%~69%)。脊椎动物如七鳃鳗、鲤鱼、乌龟、鸽子和马的细胞色素C序列,与无脊椎动物蚕蛾的细胞色素C差异程度都相近(27%~30%)。比较鲤鱼和牛蛙、海龟、鸡、兔、马的细胞色素C,其差异均为13%~14%。
也就是说,没有中间过渡类型的细胞色素C,这一点,与第一章提到的没有中间过渡物种类似,再一次否定了达尔文所提出的物种从低到高慢慢进化的错误假设。
这些分析表明,如果进化树的概念是存在的,而且细胞色素C基因的变化可以代表整个基因组的变化的话,应该出现从圆口纲脊椎动物→鱼→两栖动物→爬行动物→哺乳动物或鸟按照进化关系远近成比例的细胞色素C基因的差异,但是并没有这些证据,反而证明了各种物种的产生应该是突然爆发的,基本是平级的关系,就像线粒体DNA条形码研究分析的结果那样。
总之,无论用哪一个“分子钟”,都会出现与“进化假说”理论发生矛盾的结果,不能自圆其说。
也就是说,即使用“进化树”“分子钟”的理论来计算,还是推导出与“进化树”自相矛盾的结果,有力地证明了这个所谓的理论体系根本就是错的。
七、基因的未解之谜
除了上述的局限性,目前所采用的基因都是编码蛋白质的基因,而其实生物体内还有大量的非编码基因,这些基因在“系统进化树”中的位置,甚至根本没有体现出来。
7.1 基因里的“黑匣子”
人们曾经认为,人类的98%的基因都是由毫无意义的非编码、无功能的DNA组成的,又被称为“垃圾”DNA。最近的发现表明,这与事实相去甚远。²⁶³
人类基因组有大约20,000个蛋白质编码基因,其中包含制造构建、维护和修复我们身体所需的特定蛋白质的指令。这些基因仅占我们基因组的2%,长期以来,我们DNA的其余98%被认为是“垃圾”。然而,在过去十年中,研究人员越来越多地使用全基因组测序和其它“组学”技术,对非编码区域有了重新认识。
基因使用三碱基密码来指定将形成特定蛋白质的氨基酸序列。编码蛋白质的DNA密码不是直接读取的,首先细胞会在一种非常相似的RNA分子中制作一个拷贝,这个过程称为转录。RNA包含编码蛋白质的外显子区域以及被称为内含子的非编码区域。
然后,内含子被移除,外显子被“拼接”在一起形成mRNA(信使RNA),这需要剪接体(spliceosome)的精密控制,在正确的位置将内含子切掉并将外显子连接在一起。这必须是在正确的方向和位置,如果外显子连错一个核苷酸,就会产生巨大的差异。最后,被剪接后的RNA分子在核糖体的帮助下,被解码合成蛋白质。
如果进化论是正确的,自然选择为什么会让那些被认为是垃圾的98%的基因存留下来?为什么生物会进化出如此精密复杂的内含子序列,还有如此复杂的剪切步骤才能拼接出DNA中的有编码的区域?这个过程似乎浪费了大量的时间。
认为内含子无用的认识,其实是荒谬的。人们不知其功能,并不意味着内含子没功能。
这种非编码DNA已被发现有许多用途,有充分的证据表明,它作为精心设计的遗传信息网络的一部分,具有重要作用。例如对整体基因组结构和功能进行调控。一些被称为microRNA(miRNA)的非编码RNA可以调节其它基因编码的蛋白质的产生,并且在人类、小鼠和斑马鱼体内的功能几乎相同。所以,把它当作垃圾DNA,是错误的。
7.2 蕨类植物撼动进化树
根据“进化树”的假说,进化树上较早出现的物种和较晚期才出现的物种相比,遗传物质应该更加简单、数量应该更少。可是人类有23对染色体,鸟类通常有40对染色体,更令人惊讶的是,起源于非常久远年代的一种小小的蕨类植物,竟然拥有现有地球生物中最多的染色体──720对。水蕨的基因组具有123亿个碱基,是人类基因组的3.8倍多²⁶⁴。
如果进化假说成立的话,生命应该由简单向复杂进化,为什么古代的生物会有这么多的染色体,而地球上具备最高等智慧的生物──人类,所拥有的染色体和基因数量还不如古代的一种植物呢?外表简单、基因却如此复杂的蕨类植物,显然无法用进化假说来解释。
难怪分子遗传学家罗伯特‧费尔德曼(Robert Feldman)认为:“我认为‘生命之树’的这三大区域(古细菌、细菌和真核生物)是否会站得住脚,是一个悬而未决的问题。”²⁶⁵
所以达尔文进化假说根本无法解释物种在基因上的巨大差异;而物种基因的差异,更无法推出达尔文所假定的亲缘关系。达尔文进化假说再一次被证明是错误的。
进化树是根本就不存在的“假想之树”,分子钟也是根本不存在的“假想之钟”。
因为进化论长期的影响,人们已习惯被灌输“系统进化树”所描述的生物起源假说,这样的思维模式已深入人心。若否定了系统发生树这样的理论,可能会让许多人难以接受,特别是在分子生物学领域工作的人,他们会说:“我们每天都在进行基因测序和分析基因突变的相关研究。如果一下子没有了系统发生树,那么一切物种又是从哪里来的呢?我们又该干什么呢?”
其实,所有事物都有起源,就像宇宙也有其诞生过程一样,万物也都有一个起源和建立的过程。大家都知道乐高(LEGO)玩具,数百个相似的零件,可以根据组装者的设计意图组合成许多不同的形状,每次组装出的产品都可以千差万别、截然不同,或是飞机,或是城堡等等,林林总总。
那么这些飞机和城堡是否一定需要一个共同祖先、不同组装产品之间是否一定需要一个亲缘关系呢?实际上,进化论起源于无神论的思考方式,仅仅基于对物质世界的表面观察,并套用了错误的理论和分析方法,得出了错误的结论的同时,使人加深了对无神论的盲目膜拜、放弃了对生命的精神和灵魂层面的探索。
既然我们已经发现乘坐的是一艘漏洞百出的破船,何不干脆放弃、另外找一个更安全牢固的船舶顺利到达目的地?没有了“系统进化树”、没有了进化论,地球照样运转,物种照样繁荣。放下了深重思想枷锁的人,可能更有时间、更有机会来深入思考生命现象的本质,探索生命和大千世界的奥秘。在接下来的章节中,我们将从更多层面对此进一步阐述。
八、否定进化假说的十三个要点
无论从理论到实验、从宏观到微观、从历史到现代、从细胞到分子、从传统和现代生物学的角度,无论何时何地、用何种方法透视进化假说,人们都没有找到任何支持进化假说的证据;相反,下面十三个方面的错误,足以否定进化论,总结如下:
第一,以偏概全:达尔文一开始就犯了基本逻辑错误,用种内变异(后被称为“微进化”)的例子错误地推导出种间进化(后被称为“广进化”)的结论。
第二,混淆概念:用“微进化”鱼目混珠、模糊其与“宏进化”之间的巨大差异,达到混淆视听的效果。
第三,验证失败:科学家做了各种各样的实验,包括米勒烧瓶实验、乔伊斯等做的RNA实验和兰斯基的大肠杆菌实验,试图验证进化假说,结果都以失败告终。
第四,违反自然规律:亘古不变的“活化石”和“人工育种”不产生新物种等基本科学事实,都证明环境不能改变物种。达尔文的进化思想违反了自然界物种保持稳定的铁律。
第五,人不是猴子变的:猴子和人之间,从生物学、基因、蛋白质、社会学和思想行为方面来看都差异巨大,本质完全不同;没有化石证明人猿先祖的存在,没有任何证据能证明或推导出猴子可能进化成人。
第六,不是“慢慢演化”而是“突然爆发”:没有一块化石能证明进化假说,寒武纪化石和线粒体DNA研究提示地球上的生命是以大爆发的形式产生的;地球上的生命不是单一线性慢慢演化,更可能是多次爆发→灭绝的重复循环。
第七,不是“自然选择”而是“有意设计”:长颈鹿的脖子为什么这么长?“进化论”所解释的食物短缺和性选择假说都是牵强附会的无稽之谈;达尔文在《物种起源》中坦率承认“如果以为眼睛是可以通过进化形成的,那就真是荒诞到极点”。
第八,看似“结构退化”其实“大有用处”:人体功能十分复杂精妙,不是人眼所看到的大小结构反映出来的,结构不能代表功能,结构变小不代表功能上变弱,例如切除扁桃体可能会增加感染、肺病等发病风险;松果体虽然萎缩,但对内分泌、神经、视觉、生殖和免疫都发挥着重要作用;胸腺虽然萎缩,却还具备调节内分泌、神经和寿命的功能。
第九,不是“残酷竞争”而是“共生互惠”:尽管在自然界中也存在竞争和求生的压力,但达尔文错在把它绝对化、极端化地描述成为一种规律,用这种弱肉强食、残酷竞争的冷酷关系,定义自然生物之间的基本关系,以此作为“物种进化”的动力,而其实,自然界生物之间能够稳定繁荣下去的基本关系是“共生互惠”,共生互惠的基本关系有助于维持物种的多样性和生态的稳定性。生物界中“共生互惠”的例子俯拾皆是,已有科学实验证明老鼠和猩猩都自发地帮助弱势同伴,何况有理性、有道德的人?有些人把基因说成是“自私”的,但基因本身并没有道德或善恶的概念,人作为有理性和道德的生物,可以自觉地追求道德和善良。
第十,不是“科学事实”而是“欺骗造假”:在博物馆、科学论文、教科书中,后人为了支持进化假说而编造出来的假证据比比皆是。德国海克尔编造的“胚胎重演论”“皮尔敦猿人”被BBC称为英国史上最大的科学骗局,所谓的“内布拉斯加男子”原来是猪。
第十一,生命的复杂高效否定“进化”:生物系统、细胞极为复杂,且可以高效运作,绝对不可简化。且不说让达尔文感到震撼的眼睛,真核细胞就像一个最先进的笔记本电脑,肝脏细胞就像人体的生化工厂,细菌鞭毛堪称高效精密的分子马达,人体的免疫系统像是一个智慧奇妙的海陆空防御系统,神经系统像是一个复杂精妙的信息接收处理系统,这些都完全无法用进化假说进行解释。
第十二,基因与“进化”背道而驰:大量基因研究的现代成果毋庸置疑地说明大部分基因突变其实是“有害”的,生命主旋律不是“进化”,而是“退化”。中性学说从根本上动摇了自然选择假说,也就是某个基因能不能被保留下来,不是靠环境选择,而是靠某种不可知的“运气”;“定向突变”也对达尔文进化假说体系的“随机突变”理论形成巨大挑战。
第十三,“系统发生树”是没有根据的假想之树:树上的“节点”是假想的,生物界不同物种之间的亲缘关系是假想的;在系统进化树的分析中会出现不可避免的拓扑学错误。用达尔文错误的“物种进化”思想试图去理解生命的起源、解释生命基因的大数据,就像盲人摸象,永远也无法获知大象的全貌,又像海中捞月,永远只能是幻想。
开始拿到基因数据的科学家如获至宝,可是难题纷沓而来:到底选哪个基因?同样数据,用不同的方法分类,会产生很不一样的辈分关系。刚刚还是爷爷,又被鉴定为孙子,辈分混乱;不同速率滴答作响的分子钟也让人无所适从,无论用哪个分子钟,都能推导出自相矛盾的结果,这一点足以证明“进化树”是一个完全错误的理论体系;基因里的“黑匣子”非编码DNA,更是让科学家无从下手;一种小小的蕨类植物,居然拥有地球生物中最多的染色体,这也足以撼动进化树。
总而言之,进化假说没有任何科学凭据,本质上违反基本科学精神、违反科学原理。进化假说是伪科学。
否定进化假说的十三个要点。(大纪元)
九、千名科学家质疑“20世纪最大的谎言”
从1859年达尔文的《物种起源》问世以来,不断受到宇宙学、物理学、生物学等多学科和新发现的挑战。达尔文的进化假说与事实之间有很多矛盾。随着时间的推移和研究的深入,这些矛盾变得更加尖锐、不可调和。
分子生物学家、医学博士道顿(Michael Denton)在1985年出版的《进化论:陷入危机的理论》(Evolution: A Theory in Crisis)一书中称达尔文进化假说是“20世纪最大的谎言”。²⁶⁶
2001年美国一群优秀科学家创立了美国智库发现研究所(Discovery Institute),他们在发起的“对达尔文主义进化假说的科学质疑”宣言中写道²⁶⁷:“我们质疑达尔文宣称的用生物随机突变和自然选择论来解释复杂的生命,我们鼓励(人们)谨慎检验达尔文理论存在的证据。”
宣言联署者来自美国、英国、日本、俄国、以色列、香港、捷克、匈牙利、印度、尼日利亚、波兰等多个国家和地区,是从事进化生物学、生物学、化学和其它自然科学研究的国际著名教授、博士、科学院院士等。他们中有些在国际知名大学从事科研工作,如英国剑桥大学、美国哈佛大学、耶鲁大学、哥伦比亚大学、斯坦福大学、加州大学、康奈尔大学、普林斯顿大学、布朗大学、宾夕法尼亚大学等。
联署者还包括多名曾得到诺贝尔奖提名的科学家,如进化生物学家士丹利‧萨尓斯(Stanley Salthe)、乔治亚大学量子化学家亨利‧舍费尔(Henry Schaefer)、俄罗斯自然科学院胚胎学家利瓦伊‧比洛斯夫(Lev Beloussov)和美国科学促进会研究员赖尓‧詹森(Lyle Jensen)。
“发现研究所”的网站上写道:“这是科学家关于他们──对与新达尔文主义有关的科学证据的评估以及对现代达尔文理论证据有仔细审查的必要性──给予肯定的专业声明。”“达尔文主义的科学异议声明是为了纠正公共记录,表明有科学家支持公开审查与现代达尔文理论有关的证据,并质疑新达尔文主义能否解释自然世界的复杂性和多样性。”
截至2020年4月,已经有化学、生物学、医学、物理学、地质学、人类学、古生物学、统计学及其它领域的一千一百多名科学家和研究人员签署了这份针对达尔文进化假说的质疑宣言。
2018年,印度人力资源发展部部长萨蒂亚帕尔‧辛格(Satyapal Singh)认为达尔文理论“从科学上来说是错误的”,因为没有“看到任何一只猴变人”,并敦促将达尔文进化论从学校课程中删掉²⁶⁸。印度新德里的一些学校已经放弃教授达尔文的进化假说²⁶⁹。
十、为什么进化假说一直持续至今?
尽管科学家们发现进化假说是一个漏洞百出的学说,为什么它在科学界仍然占据主导地位、欺骗世界一百多年?其中原因众多。
首先,科学界具有的思维惯性。现代科学的发展通常是以前人的研究为基础,吸收其成果,通常不太会彻底抛弃或推翻之前的研究。因为理论学说的惯性,一些原有理论体系的内容已经被发现是错的,但因为其中有些观点看上去似乎也有一定道理,学术界还是把它作为一种研究或观点保留,不彻底推翻。科学界的思维惯性一般都不愿意跳出原来的思维框架去彻底反思。
第二,进化假说片面认为人是物质结构的组合体,忽视了人的能量和精神特性,而后者才是人更本质的属性。停留在单纯的物质层面上探索生命起源,不可能抓其本质、看到真相,只能被进化假说越绕越糊涂。这一点会在第四、五章详细论述。
第三,未经证明的进化假说,已经堂而皇之地进入学校课堂被大肆宣讲。质疑“进化假说”的科学发现被歪曲,质疑的文章被延误发表,质疑“进化假说”的科学家被打压。这种学术观点的排他性垄断,实际上对人们起到了某种程度的洗脑作用,造成越来越多的危害、形成恶性循环而不能自拔。这一点会在第六章详细论述。
第四,以分析型思维为主的现代实证科学,试图寻找一种和传统文化不同甚至对立的对生命起源的解释。这种从未证实的假说却被扩展到了社会科学和人文科学中,造成的恶果远远超出生物学领域。进化假说的生存竞争逻辑一旦运用于社会实践,就摧毁了人类道德的根基,使人类物化以致堕落。这一点会在第七章详细论述。
第五,19世纪宗教败坏后,人类社会出现信仰危机,加上工业革命后科学技术蓬勃发展、人类生产力大增、可以使用机械,进化假说是在这样的背景下产生的。就如青少年进入叛逆期一样,人们容易抛弃传统的价值观,寻求新奇和叛逆的学说。这一点我们会在第八章详细论述。
前面的第一章,我们为大家疏理了进化假说的逻辑错误、否定进化论的实验数据和科学事实;第二章是破除进化假说的基本迷思;第三章深入解读了否定进化假说的现代分子生物学研究成果。
基于前面三章的大量科学研究和试验证明,进化假说已经被彻底否定了。它甚至连假说都谈不上,更应该从教科书中被删除。那么,人的生命到底是怎么来的,我们在下面的章节将继续探讨。待续。△
参考文献:
230. Baum, D. (2008) Reading a Phylogenetic Tree: The Meaning of Monophyletic Groups. Nature Education 1(1):190.
https://tinyurl.com/yyfy9nay
231. By Ernst Haeckel – First version from en.wikipedia; description page was here. Later versions derived from this scan, from the American Philosophical Society Museum., Public Domain, https://commons.wikimedia.org/w/index.php?curid=2731459
232. Willi Hennig. Phylogenetic Systematics. Translated by D. Dwight Davis and Rainer Zangerl. University of Illinois Press Urbana and Chicago. 1966.
https://tinyurl.com/35u969hf
233. Woese, C. R., Kandler, O., & Wheelis, M. L. (1990). Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. Proceedings of the National Academy of Sciences of the United States of America, 87(12), 4576–4579.
https://doi.org/10.1073/pnas.87.12.4576
234. Woese, C. R., & Fox, G. E. (1977). Phylogenetic structure of the prokaryotic domain: The primary kingdoms. Proceedings of the National Academy of Sciences, 74(11), 5088-5090.
https://doi.org/10.1073/pnas.74.11.5088
235. Hedges, S. B., Marin, J., Suleski, M., Paymer, M., & Kumar, S. (2015). Tree of Life Reveals Clock-Like Speciation and Diversification. Molecular Biology and Evolution, 32(4), 835-845.
https://doi.org/10.1093/molbev/msv037
236. Hinchliff, C. E., Smith, S. A., Allman, J. F., Burleigh, J. G., Chaudhary, R., Coghill, L. M., Crandall, K. A., Deng, J., Drew, B. T., Gazis, R., Gude, K., Hibbett, D. S., Katz, L. A., Laughinghouse, H. D., McTavish, E. J., Midford, P. E., Owen, C. L., Ree, R. H., Rees, J. A., . . . Cranston, K. A. (2015). Synthesis of phylogeny and taxonomy into a comprehensive tree of life. Proceedings of the National Academy of Sciences, 112(41), 12764-12769.
https://doi.org/10.1073/pnas.1423041112
237. Hug, L. A., Baker, B. J., Anantharaman, K., Brown, C. T., Probst, A. J., Castelle, C. J., Butterfield, C. N., Hernsdorf, A. W., Amano, Y., Ise, K., Suzuki, Y., Dudek, N., Relman, D. A., Finstad, K. M., Amundson, R., Thomas, B. C., & Banfield, J. F. (2016). A new view of the tree of life. Nature Microbiology, 1(5), 1-6. https://doi.org/10.1038/nmicrobiol.2016.48
238. Kumar, S., Suleski, M., Craig, J. M., Kasprowicz, A. E., Sanderford, M., Li, M., Stecher, G., & Hedges, S. B. (2022). TimeTree 5: An Expanded Resource for Species Divergence Times. Molecular Biology and Evolution, 39(8).
https://doi.org/10.1093/molbev/msac174
239. Hall, B. G. (2013). Building Phylogenetic Trees from Molecular Data with MEGA. Molecular Biology and Evolution, 30(5), 1229-1235.
https://doi.org/10.1093/molbev/mst012
240. Cambrian Period. National Geographic.
https://tinyurl.com/8fufk73y
241. M.Y. Stoeckle, D.S. Thaler. Why should mitochondria define species? Human Evolution. Vol. 33; n. 1-2 (1-30) – 2018.
DOI: 10.14673/HE2018121037; https://tinyurl.com/yc28xwbh
242. Amy Joi O’Donoghue. Why this 300 million-year-old fossil discovered in Utah has the paleontology world buzzing? Rare creature found in Utah’s Canyonlands National Park. Nov 8, 2021, 8:24pm CEST.
https://tinyurl.com/3snhan29
243. McNutt, E. J., Hatala, K. G., Miller, C., Adams, J., Casana, J., Deane, A. S., Dominy, N. J., Fabian, K., Fannin, L. D., Gaughan, S., Gill, S. V., Gurtu, J., Gustafson, E., Hill, A. C., Johnson, C., Kallindo, S., Kilham, B., Kilham, P., Kim, E., … DeSilva, J. M. (2021). Footprint evidence of early hominin locomotor diversity at Laetoli, Tanzania. Nature, 600(7889), 468-471.
https://doi.org/10.1038/s41586-021-04187-7
244. Britannica, The Editors of Encyclopaedia. “octopus”. Encyclopedia Britannica, 20 Jun. 2023, https://tinyurl.com/mr3s7fva. Accessed 17 July 2023.
245. Andrea Michelson. Octopus Arms Have Minds of Their Own.
https://www.discovery.com/science/Octopus-arms
246. Steele, E. J., Al-Mufti, S., Augustyn, K. A., Chandrajith, R., Coghlan, J. P., Coulson, S., Ghosh, S., Gillman, M., Gorczynski, R. M., Klyce, B., Louis, G., Mahanama, K., Oliver, K. R., Padron, J., Qu, J., Schuster, J. A., Smith, W., Snyder, D. P., Steele, J. A., . . . Liu, Y. (2018). Cause of Cambrian Explosion – Terrestrial or Cosmic? Progress in Biophysics and Molecular Biology, 136, 3-23.
https://doi.org/10.1016/j.pbiomolbio.2018.03.004
247. Steele, E. J., Al-Mufti, S., Augustyn, K. A., Chandrajith, R., Coghlan, J. P., Coulson, S., Ghosh, S., Gillman, M., Gorczynski, R. M., Klyce, B., Louis, G., Mahanama, K., Oliver, K. R., Padron, J., Qu, J., Schuster, J. A., Smith, W., Snyder, D. P., Steele, J. A., …Liu, Y. (2018). Cause of Cambrian Explosion – Terrestrial or Cosmic? Progress in Biophysics and Molecular Biology, 136, 3-23.
https://doi.org/10.1016/j.pbiomolbio.2018.03.004
248. Jombart, T., Archer, F., Schliep, K., Kamvar, Z., Harris, R., Paradis, E., Goudet, J., & Lapp, H. (2017). apex: phylogenetics with multiple genes. Molecular ecology resources, 17(1), 19–26.
https://doi.org/10.1111/1755-0998.12567
249. Lynch M. (1989). PHYLOGENETIC HYPOTHESES UNDER THE ASSUMPTION OF NEUTRAL QUANTITATIVE-GENETIC VARIATION. Evolution; international journal of organic evolution, 43(1), 1–17.
https://tinyurl.com/yv7zads5
250. Pennisi E. (1998). Genome data shake tree of life. Science (New York, N.Y.), 280(5364), 672–674.
https://doi.org/10.1126/science.280.5364.672
251. Pennisi E. (1998). Genome data shake tree of life. Science (New York, N.Y.), 280(5364), 672–674.
https://doi.org/10.1126/science.280.5364.672
252. Woese, C. R., & Fox, G. E. (1977). Phylogenetic structure of the prokaryotic domain: The primary kingdoms. Proceedings of the National Academy of Sciences, 74(11), 5088-5090.
https://doi.org/10.1073/pnas.74.11.5088
253. Pennisi E. (1998). Genome data shake tree of life. Science (New York, N.Y.), 280(5364), 672–674.
https://doi.org/10.1126/science.280.5364.672
254. 王博群、吴宗贤、赵淑妙。正本溯源──生物亲缘演化树的脉理和论战。中央研究院生物多样性研究中心。
https://tinyurl.com/mrxy4krh
255. Jombart, T., Archer, F., Schliep, K., Kamvar, Z., Harris, R., Paradis, E., Goudet, J., & Lapp, H. (2017). apex: phylogenetics with multiple genes. Molecular ecology resources, 17(1), 19–26.
https://doi.org/10.1111/1755-0998.12567
256. Westram, A., & Johannesson, K. (2016). Parallel Speciation. Encyclopedia of Evolutionary Biology, 212-219.
https://tinyurl.com/58md4za4
257. Gautam, P. (2020). Divergent Evolution. In: Vonk, J., Shackelford, T. (eds) Encyclopedia of Animal Cognition and Behavior. Springer, Cham.
https://tinyurl.com/d4th43zw
258. Gabora, L. (2013). Convergent Evolution. Brenner’s Encyclopedia of Genetics (Second Edition), 178-180.
https://tinyurl.com/mrkct2jn
259. Ho, S. (2008) The molecular clock and estimating species divergence. Nature Education 1(1):168.
https://tinyurl.com/27ab6j6u
260. Dr. Michael Denton, Evolution: A Theory in Crisis, Published by Adler & Adler, Distributed by Woodbine House,1985.
https://tinyurl.com/mt6rczfb
261. W. Ho, S. Y., & Duchêne, S. (2014). Molecular-clock methods for estimating evolutionary rates and timescales. Molecular Ecology, 23(24), 5947-5965.
https://doi.org/10.1111/mec.12953
262. Dr. Michael Denton, Evolution: A Theory in Crisis, Published by Adler & Adler, Distributed by Woodbine House,1985.
https://tinyurl.com/mt6rczfb
263. Alok Jha, science correspondent Wed 5 Sep 2012 20.03 BST. Breakthrough study overturns theory of ‘junk DNA’ in genome.
https://tinyurl.com/mryv4htt
264. Marchant, D.B., Chen, G., Cai, S. et al. Dynamic genome evolution in a model fern. Nat. Plants 8, 1038–1051 (2022).
https://doi.org/10.1038/s41477-022-01226-7
265. Pennisi E. (1998). Genome data shake tree of life. Science (New York, N.Y.), 280(5364), 672–674.
https://doi.org/10.1126/science.280.5364.672
266. Dr. Michael Denton, Evolution: A Theory in Crisis, Published by Adler & Adler, Distributed by Woodbine House,1985.
https://tinyurl.com/mt6rczfb
267. A Scientific Dissent from Darwinism. (2023, January 23). In Wikipedia.
https://tinyurl.com/27faup7a
268. Darwin’s theory wrong, nobody saw ape turning into man: Minister Satyapal Singh. Press Trust of India, Aurangabad | ByPress Trust of India. Jan 21, 2018.
269. John Reed and Jyotsha Singh. New Delhi. New Delhi erases Darwin from textbooks.
https://tinyurl.com/ytjkufpd
《透视“进化论”》写作组
点阅【透视“进化论”】系列文章。
前言 第一章:进化论是未经证实的假说(上)第一章:进化论是未经证实的假说(下)第二章:破除进化假说的经典迷思(上)第二章:破除进化假说的经典迷思(中)第二章:破除进化假说的经典迷思(下)第三章:分子生物学否定进化假说(上)第三章:分子生物学否定进化假说(中)





































 打印机版
打印机版


